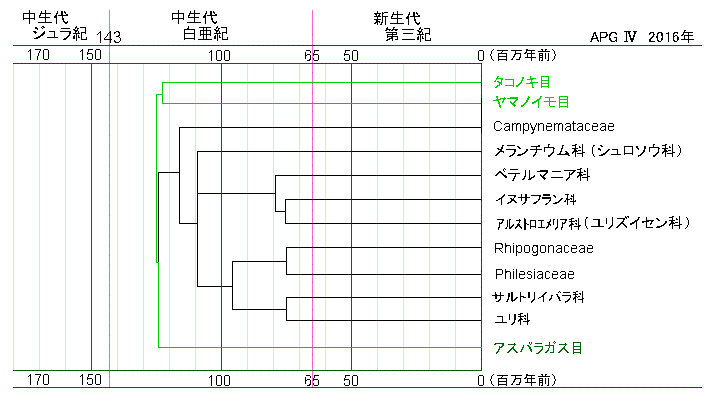
| f.07 | ユリ目の系統図 | |
 |
||
第6回 網状脈から平行脈へ
| ユリ目は、今からおよそ1億1,300万年前、タコノキ目から分かれたとされています。ちょうど、ティラノサウルスが走り回っていた白亜紀の中頃のことです。 | ユリ目には、ユリ科以外にも、コルチカムで知られるイヌサフラン科やアルストロエメリア科、サルトリイバラ科などがありますが、やはり、ユリ科が他を圧倒しています。 |
| f.07 | ユリ目の系統図 | |
|
||
エンレイソウ
| 学名 Trillium smallii Maxim. 中核単子葉植物 ユリ目 メランチウム科 (シュロソウ科) エンレイソウ属 | |
 |
エンレイソウはメランチウム科(シュロソウ科)に属します。 ユリ科が属するグループとほぼ並行に進化したグループです。 エンレイソウは、今からおよそ6,800万年前に出現しています。 大きな3枚の葉が特徴です。3枚の葉が同じ位置から出ているから、このような葉のつきかたを輪生といいます。 単子葉植物は平行脈をもつといわれていますが、エンレイソウの葉脈はどうなんでしょう。 基本的には、ヤマノイモ目の葉とよく似ていますが、3脈から横にもはっきりわかる網の目状の脈が出ているので、ヤマノイモより網状脈に近く、判断に迷います。 |
| 画像は、ホームページ「下を向いて歩こう」のmotoさんからお借りしました。 | |
 この写真もmotoさんからお借りしました。 この写真もmotoさんからお借りしました。 |
エンレイソウの花は、花弁がありません。紫色を帯びた緑褐色のがく片が3枚あります。 がく片はユリの葉のような平行脈があり、シュートのなごりを見せています。 このなかまには、3枚のがくと3枚の花弁をもつものがありますから、エンレイソウは、花弁が退化したのでしょう。 メランチウム科の場合、花被が3枚になったほうが合理的であるというわけでもありませんから、がく片や花弁が退化するのは、どうも、進化の大きな流れと直接は関係ないように思われます。 |
| 子房は基部被子植物と同じく上位で、花柱が3本出ています。おしべは6本です。 やはり、メランチウム科も3数性を示しています。 |
アルストロエメリア
| 学名 Alstroemeria sp. 中核単子葉植物 ユリ目 アルストロエメリア科 アルストロエメリア属 | |
 アルストロエメリアは、ほとんどが品種改良された園芸種で、一般にアルストロメリアと呼ばれています。 |
アリストロエメリアの出現は、エンレイソウよりも古く、今からおよそ1億年前のことでした。 内花被の2枚には斑模様があります。この模様は 種類がいくつもあるので、属名のあとに sp または ssp をつけるのが一般的になっています。sp. は、種、 ssp.は、亜種という意味です。 |
 |
外花被3枚で上の1枚は大きく同形。内花被3枚は、花被の先がとがっており、上の2枚がバンザイをしたように立っています。 花被の数は3数性を示しています。 放射相称のように見えますが左右相称です。ここがユリ科と異なる点の1つです。 おしべやめしべのつきかたは、これもユリ科と異なり、放射状にはならず、下の方にかたよって、先のほうが上に曲がっているのが特徴です。 これらの特徴は、ユリ科よりも進化しているアスパラガス目に近いように思われます。 |
 |
子房の位置は、花冠のつけねより下にありますから、子房下位です。 おしべは6本で、いちおう3数性を示していますが、ちょっと変ですね。 6本のおしべのうち、2本が小さな葯をもっています。 4本は大きな葯です。 |
 内花被の基部には、細かい毛が生えています。 顕微鏡で観察してみます。 |
アルストロエメリア科の植物が初めてこの世に出てきたころは、花粉を運ぶ昆虫は、チョウやハチ以外にも甲虫が活躍していました。甲虫というのはコガネムシなど、上の羽がかたくなった昆虫です。 甲虫は、花粉を運んでくれるだけならよいのですが、大切な胚珠までも食べてしまうことがあるのです。 アルストロエメリアは、大切な胚珠の入った子房を奥のほうにかくし、花の内側から奥に入れないように細かい毛をたくさん生やしたとも考えられます。 このような毛は |
 |
花冠の中をアップしてみます。 大きい この花の場合、小さい葯のおしべは2本でした。 他の花も調べてみると、小さい葯のおしべのほうが多いものもありました。数の比は決まっていないようです。 |
 |
小さい葯を拡大してみます。 葯室がはじけて花粉がいっぱい吹き出していました。 |
 |
大きい方の葯は、まだ、はじけていません。 どうも、大きい葯と小さい葯のはじける時期がずれているようです。 左の写真は、葯の外側の表面です。 右の写真は、葯室のさけめの中を見たものです。花粉がたくさんつまっています。 |
 |
|
| 小さい葯の花粉です。 | 大きい葯の花粉です。 |
| 小さいほうの葯の花粉のほうが、いくぶん細長いようです。 | |
 |
| アルストロエメリアのめしべです。 赤い部分が おしべに比べてかなり短いようです。 自家受粉をしにくくしているのでしょうか。 |
 |
 |
| めしべの先端、柱頭が閉じた状態です。 | 正面から見てみます・ |
 |
柱頭をむりやりに開いてみました。柱頭の内側にはたくさんの突起が見られます。 これは、どう見てもユリ科よりは進化しているように思えます。 系統図を見ると、ユリ科は、アルストロエメリア科よりだいぶ下の方にあります。下ほど進化していると見なされていますが、枝分かれしたそれぞれの科の中でもさらに進化は進んでいるはずです。 アルストロエメリア科の中では、ボマレア属が古く、アルストロエメリア属はこの科の中では新しい方なので、進化が進んでいるということになるのでしょう。 |
 |
| 柱頭の内側です。 たくさんの突起がひしめき合っています。 ここで受粉するのです。 右上に見えるひときわ大きなものは、花粉ですね。 |
 |
子房の近くで花柱を切断しました。 切り口は、ご覧のように△です。 めしべの進化は、何本かのめしべが合生して1本になるのがふつうですが、花柱は、たいてい円柱形になります。 めしべは、柱頭-花柱-子房とひと続きになって、1個の心皮となります。 ユリ目では、3個の心皮の合成がヤマノイモ目ほど単純ではないにしても、外から見てわかる程度に密着したものといえるでしょう。 |
 |
今度は、子房を輪切りにしてみました。 単子葉植物は、どれも中軸胎座です。 アルストロエメリアも例外ではありませんでした。 3個の心皮が3室をつくっています。1室には2列に胚珠がつきます。 |